Teste final mostra que não havia aftosa no rebanho dos EUA
13 de outubro de 2003Brasil retoma importação de carne argentina
15 de outubro de 2003Sobrevivência no Fluído Ruminal de Bactérias Láticas de Inoculantes de Silagem
Luiz Gustavo Nussio1 e Carla Maris Bittar Nussio2
Introdução
Inoculantes contendo principalmente bactérias láticas (BAL) são utilizados como aditivos de silagem a fim de melhorar a eficiência de sua preservação. Dentre as BAL, as utilizadas com mais freqüência são as espécies homofermentativas como Lactobacillus plantarum, Enterecoccus faecium e Pediococcus spp. Estas espécies são utilizadas devido a sua eficiente utilização de carboidratos solúveis presente nas plantas, intensiva produção de ácido lático e rápida redução no pH do material ensilado (Weinberg & Muck, 1996). Outra espécie de BAL utilizada é a Lactobacillus buchneri, a qual apresenta alta produção de ácido acético na silagem, inibindo o desenvolvimento de fungos e, portanto, preservando silagens suscetíveis a deterioração aeróbia (Weinberg et al., 2002).
Conforme foi apresentado em artigos anteriores, em alguns casos, o efeito do uso do inoculante em silagem pode ser também observado no desempenho de animais. O consumo de alimentos, o ganho de peso vivo, a eficiência alimentar e/ou a produção de leite foram melhorados entre 5-11% em 25-40% dos estudos revisados por Muck (1993).
A causa para o melhor desempenho de animais ainda não está clara. No trabalho realizado por Keady & Steen, 1995, quando o inoculante LAB foi adicionado à silagem imediatamente antes de seu fornecimento aos animais, não foram observados efeitos na MS, fração N, FDN, ou digestibilidade da FDN . Estes dados sugerem que os benefícios em desempenho de animais se devem à fermentação da silagem e não aos efeitos diretos destes inoculantes no rúmen. Entretanto, em estudo recente, a adição de BAL na dieta aumentou o consumo de MS, o ganho de peso e a digestibilidade de MS em bezerros (Khuntia & Chaudhary, 2002). Outros trabalhos também demonstraram benefícios da adição de inoculantes aplicados diretamente na dieta de ruminantes (Salawu et al., 2001; Malik & Sharma, 1998).
Os resultados de alguns desses estudos sugerem um possível efeito probiótico de BAL utilizadas como inoculantes para silagens. Entretanto, o mecanismo deste efeito ainda não é claro. Uma hipótese é que linhagens específicas de BAL possam interagir com microrganismos de rúmen, melhorando sua funcionalidade e, consequentemente, o desempenho de animais. Outra hipótese é que as BAL utilizadas como inoculantes de silagem possam inibir microrganismos prejudiciais à silagem. Já se sabe que as BAL produzem uma série de substâncias antimicrobianas, como as bacteriocinas (Muller et al., 1996).
A não ser que os efeitos benéficos desses inoculantes se deva à produção de bacteriocinas, as BAL ingeridas por animais deveriam sobreviver no fluído ruminal para que apresentassem efeito. Este trabalho teve como objetivo, avaliar a sobrevivência no fluído ruminal de BAL de inoculantes comerciais para silagem.
Material e Métodos
Fluído ruminal foi coletado de 2 vacas Holandesas portadoras de cânulas ruminais, recebendo dieta total à base de silagem de alfafa, silagem de milho e concentrado contendo milho e farelo de soja.
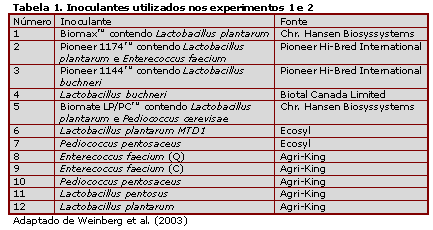
O fluido ruminal foi filtrado através de pano de queijo, sendo dividido em vários frascos estéreis, os quais foram inoculados com inoculantes comerciais (Tabela 1) contendo BAL (107 UFC/mL). Fluido ruminal sem inoculante foi utilizado como controle. Os frascos contendo fluido ruminal inoculados foram então subdivididos em dois, sendo adicionada solução de glicose em um dos frascos. Aos vários frascos, foi aplicado CO2 antes de seu fechamento e incubação a 39ºC. Amostras para análise foram coletadas após 6, 12, 24, 48 e 72h após a inoculação.
A contagem de BAL, assim como a determinação das concentrações de ácidos lático e ác. graxos voláteis (AGV) foi determinada.
Resultados
Os inoculantes foram testados em 2 experimentos: experimento 1, utilizando-se os inoculantes 1 a 5; experimento 2, utilizando-se os inoculantes 6 a 12. O fluído ruminal fresco antes da inoculação apresentou contagem de BAL de 6,2 e 5,7 log10 UFC/mL e pH de 5,7 e 5,57, para os exp. 1 e 2, respectivamente. Portanto, a inoculação resultou em um aumento de aproximadamente 10 vezes no número de BAL.
A Tabela 2 apresenta os resultados de pH final e número final de BAL. No fluído ruminal sem adição de glicose, o pH final foi numericamente superior aos valores do fluído ruminal fresco (0,07 a 0,31 unidades). Com adição de glicose, houve um decréscimo do pH comparado aos valores do fluído fresco, variando de 0,30 a 0,52 unidades. O pH final foi significativamente afetado (P<0,05) pela adição de glicose no exp. 2 e pelos inoculantes utilizados no exp. 1. A contagem final de BAL esteve em torno de 1 unidade logarítmica dos valores iniciais, com exceção do controle sem adição de glicose no exp.2, onde as contagens aumentaram aproximadamente 1,5. Todavia, a adição de glicose aumentou significativamente a contagem de BAL, mas a adição do inoculante não afetou esta contagem em ambos os experimentos. A interação entre glicose e o inoculante foi significativa no exp. 2 e, foram observadas (Tabela 2) diferenças significativas entre a contagem de BAL de diferentes inoculantes, com e sem a adição de glicose. Os inoculantes que resultaram em menores valores de pH tenderam a apresentar as maiores contagens de BAL; e aqueles que resultaram em maiores pH também tenderam a ter as menores contagens de BAL, independentemente da adição de glicose.

Nos exp. 1 e 2, a adição de glicose resultou em menores valores de pH durante o período de incubação, quando comparado com a não adição de glicose. Sem glicose, os inoculantes 4 e 5 resultaram em tamponamento similar ao inoculante 1, e os inoculantes 2 e 3 foram similar. Sem glicose, a maior parte dos inoculantes desempenhou como o inoculante 8 no exp. 2; o inoculante 7 teve um nível intermediário de tamponamento, comparado aos inoculantes 6 e 8. Com glicose, a mudança de pH durante o período de incubação foi similar àquela do inoculante 1 no exp.1, e todos os inoculantes do exp. 2 apresentaram padrão de mudança de pH similar.
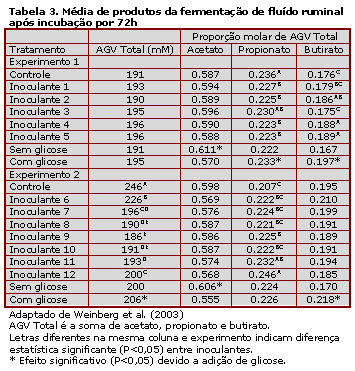
O fluído ruminal fresco apresentou concentração de 97-98 mM de acetato, 35-38 mM de propionato e 22-24 mM de butirato, concentrações típicas observadas no rúmen. A concentração de AGV na amostra controle aumentou durante a incubação e alcançou valores de 110-148 mM de acetato, 43-54 mM de propionato e 32-49 mM de butirato, após 72h de incubação. As concentrações no fluído ruminal contendo inoculante foram: 105-139 mM de acetato, 40-52 mM de propionato e 28-52 mM de butirato. Não foi possível detectar lactato nas amostras analisadas. Baixas concentrações de etanol (1-1,4 mM) foram detectadas nas amostras contendo inoculante e glicose no exp.1, com exceção do inoculante 2.
A Tabela 3 apresenta a concentração média de AGV nas 72h de incubação dos diferentes tratamentos. Foram observados efeitos significativos do inoculante e/ou da adição de glicose. As únicas interações inoculante-glicose significativas foram observadas para a concentração de butirato no exp.1 e AGV total no exp.2. No exp. 1, as concentrações de propionato e butirato foram significativamente afetadas. No exp.2, no qual as concentrações de AGV nas amostras controle foram altas, a inoculação suprimiu a formação destes ácidos. A inoculação no exp.2 aumentou a fração molar e houve consistente tendência de redução de acetato. Em ambos os experimentos, a adição de glicose reduziu a formação de acetato e aumentou a de propionato e butirato.
Discussão
A taxa de inoculação do fluído ruminal fresco (107 UFC/mL) foi determinada para ser comparável ao número de BAL consumidas por vacas que recebem 45 kg de silagem (peso fresco) em sua dieta diariamente. As BAL não são importantes para rúmen de animais alimentados basicamente com forragens, exceto quando grandes quantidades de carboidratos solúveis ou amido estão disponíveis (Van Soest, 1994). Entretanto, os resultados deste trabalho indicam que as BAL testadas são capazes de sobreviver e, em alguns casos, de crescerem em fluído ruminal. Como esperado, a adição de glicose aumentou de forma marcante a sobrevivência das BAL no fluído ruminal, sugerindo que as espécies de BAL de inoculantes de silagem podem competir efetivamente com a microflora ruminal na presença de glicose. Algumas espécies de BAL (inoculantes 5, 6 e 9) cresceram mais que outras. O inoculante 6 (L. plantarum) resultou ainda na maior concentração de AGV no fluído ruminal, quando comparado aos outros inoculantes. Esta espécie têm sido associada aos efeitos benéficos no desempenho de animais em estudos conduzidos na Irlanda (Keady et al., 1994).
O fato de não ter sido detectado ácido lático no fluído ruminal inoculado não significa que os inoculantes não resultaram na produção deste ácido. Pode ter ocorrido rápida conversão do ácido lático produzido à AGV por microrganismos do rúmen. Geralmente as concentrações de ácidos lático são menores que aquelas observadas para AGV (Van Soest, 1994). Em relação ao controle, as amostras inoculadas apresentaram elevadas concentrações de butirato no experimento 1 e maiores concentrações de propionato no experimento 2. Estes são potenciais produtos finais da fermentação de lactato dependendo da população microbiana presente no rúmen. De forma alternativa, as BAL podem não ter fermentado a glicose mas podem ter afetado a população microbiana responsável por essa ação.
A observação de que o pH do fluído ruminal de amostras inoculadas foi maior que o controle não era esperada, uma vez que, em silagens as BAL causam rápido declínio de pH como resultados da produção de ácidos orgânicos, principalmente ácidos lático. As espécies utilizadas também variaram em sua habilidade em tamponar o pH, tanto com quanto sem a adição de glicose. Os inoculantes 3, 2 e 8 foram os melhores na manutenção do pH próximo aos seus valores iniciais. Este fenômeno sugere que o modo de ação dos inoculantes no fluído ruminal deve estar mais relacionado à sua influência nos microrganismos predominantes no rúmen, do que à direta fermentação de substratos por BAL no rúmen. Maiores valores de pH devem certamente melhorar a funcionalidade de microrganismos específicos do rúmen, especialmente quando ocorre decréscimo de pH devido ao fornecimento de alta energia (Van Soest, 1994). Ainda, o aumento na digestibilidade da fibra de silagens inoculadas já foi observado (Muck, 1993), e este efeito de tamponamento pode ser uma possível explicação, uma vez que o crescimento de bactérias fibrolíticas é inibido por pH menor que 6.
Conclusão
Os resultados deste trabalho indicam que as BAL de inoculantes podem sobreviver no fluído ruminal. Estas BAL levam a algumas mudanças no fluído ruminal tais como mudanças no pH e proporção de AGV. Como estas mudanças afetam o desempenho animal ainda não foi elucidado, havendo necessidade de mais pesquisa nesta área.
Referências
Keady, T.W.J. & Steen, W.J. The effects of treating low dry-matter, low digestibility grass with bacterial inoculant on the intake and performance of beef cattle, and studies of its mode of action. Grass and Forage Sci., v.50, p.217-226, 1995.
Khuntia, A. & Chaudhary, L.C. Performance of male crossbred calves as influenced by substitution of grain by wheat bran and the addition of lactic acid bacteria to diet. Asian-Australian J. Anim. Sci., v.15, p.188-194, 2002.
Malik, R. & Sharma, D.D. In vitro evaluation of different probiotics as feed supplement. Indian J. Dairy Sci., v.51, p.357-362, 1998.
Muck, R.E. The role of silage additives in making high quality silage. In: Silage production from seed to animal. Pp. 106-116. Syracuse, NY:NRAES-67, Northeast Regional Agricultural Engeneering Service.
Muller, T.; Behrendt, U.; Muller, M. Atagonistics activity in plant-associated lactic acid bacteria. Microbiol. Res., v.151, p.63-70, 1996.
Van Soest, P.J. Function of the ruminant forestomach. In: Nutritional Ecology of the Ruminant. Ed. Van Soest, P., pp.230-252. Ithaca, N.Y.USA: Cornell University Press.
Weinberg, Z.G. & Muck, R.E. New trends and opportunities in the development and use of inoculants for silage. FEMS Micro. Reviews, v.19, p.53-68, 1996.
Weinberg, Z.G.; Ashbell, G.; Hen, Y; Arieli, A.; Szakacs, G.; Filya, I. Ensiling whole-crop wheat and corn in large containers with Lactobacillus plantarum and Lactobacillus buchneri. J. Industrial Micro. and Biotech., v.28, p.7-11, 2002.
Weinberg, Z.G.; Muck, R.E.; Weimer, P.J. The survival of silage inoculant lactic acid bacteria in rumen fluid. J. Applied Micro., v.94, p.1066-1071, 2003.
_______________________________________________
1Prof. Dr. Depto. de Zootecnia – ESALQ/USP
2Pesquisadora Embrapa Pecuária Sudeste
0 Comments
muito bom trabalho, isso ajuda a tirar duvidas quanto a eficiencia do inoculante.