Fertilidade em novilhas após aborto induzido com cloprostenol
16 de abril de 2003Dia de campo e Leilão Paredão nos dias 25 e 26 de Abril
16 de abril de 2003Consumo alimentar em bovinos de corte
Por Rodrigo de Almeida1 e Dante Pazzanese Lanna2
O consumo de matéria seca (CMS) é um parâmetro fundamental na formulação de dietas para atender as exigências nutricionais, e estimar tanto o ganho de peso diário dos animais como a lucratividade da exploração, particularmente em confinamento. Segundo o último NRC para Bovinos de Corte (National Research Council, 1996), os fatores que regulam o CMS pelos ruminantes são complexos e não completamente conhecidos. Ainda assim, estimativas do consumo são essenciais para predizer as taxas de ganho de peso e para a aplicação das equações de predição das exigências nutricionais de gado de corte.
Pesquisas anteriores (Van Soest, 1994) estabeleceram relações entre a concentração de energia na dieta e o CMS em gado de corte, baseado no conceito de que o consumo de dietas de menor digestibilidade e de baixa energia (geralmente com altos percentuais em fibra) é controlado por fatores físicos tais como enchimento ruminal e taxa de passagem, enquanto que o consumo de dietas de alta digestibilidade e alta energia (geralmente baixa fibra e alta proporção de concentrados) é controlado pelo atendimento da demanda energética do animal e por fatores metabólicos.
Há muito tempo reconhece-se que existe significativa variação entre os animais no consumo alimentar, na exigência de manutenção e na eficiência da utilização dos alimentos (Ferrell & Jenkins, 1995). Bovinos Bos indicus puros e cruzados utilizam forragens de baixa qualidade mais eficientemente do que bovinos Bos taurus (Karue et al., 1972, e Krehbiel et al., 2000). Isto explica o superior desempenho dos cruzados Bos indicus x Bos taurus em relação aos animais Bos taurus em condições de ambiente subtropical, particularmente em ambientes com restrições nutricionais importantes (Frisch & Vercoe, 1977).
Por outro lado, quando fornecemos dietas a base de forragem de alta qualidade ou de forragem e concentrado, bovinos Bos taurus consomem mais alimento em relação às suas exigências de manutenção e assim ganham peso mais rápido e eficientemente do que animais Bos indicus (Ledger et al., 1970 e Krehbiel et al., 2000). Isto é particularmente verdadeiro em condições não limitantes de produção, como dietas de confinamento e dietas à base de pastagens com alta inclusão de concentrados.
Alguns trabalhos e dados coletados de diversos confinamentos aqui no Brasil confirmam padrões bem distintos de consumo entre zebuínos puros e europeus puros ou cruzados. Nota-se um menor consumo para zebuínos na ordem de 10 a 12%. Este menor consumo de genótipos zebuínos foi recentemente verificado por Almeida & Lanna (2003).
Nas Figuras 1 e 2 observa-se que os consumos de tourinhos das raças Angus e Brangus foram significativamente (P<0,01) maiores do que o consumo de tourinhos Nelore à mesma idade (entre 9 e 13 meses de idade). Figura 1 – Médias ajustadas de consumo (kg MS/dia) para tourinhos em prova de ganho de peso
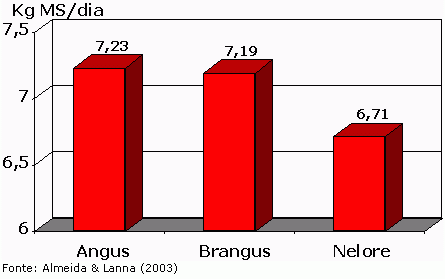
Figura 2 – Médias ajustadas de consumo (%PV) para tourinhos em prova de ganho de peso
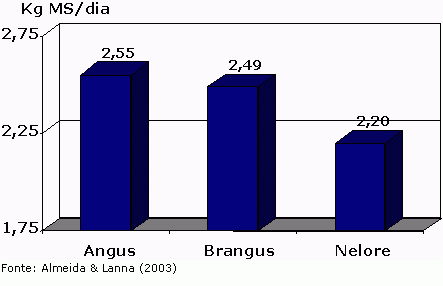
Por outro lado, quando se comparam estimativas de consumo entre bovinos europeus puros vs. bovinos cruzados (zebuínos x europeus), observam-se maiores semelhanças entre os dois genótipos na literatura consultada (por exemplo, Krehbiel et al., 2000). Esta similaridade no padrão de consumo também é bastante pronunciada na comparação entre raças européias britânicas e continentais. Entretanto, outras pesquisas demonstraram maiores consumos (Beaver et al., 1989, Boyles & Riley, 1991, Huffman et al., 1990 e Kappel et al., 1972) em bovinos europeus comparados com bovinos cruzados com Bos indicus, principalmente em dietas com alta proporção de grãos. As razões para estas pequenas discrepâncias entre experimentos podem estar relacionadas com diferenças genéticas e ambientais.
Ao mesmo peso, novilhos Holandeses consomem consideravelmente mais alimento que novilhos de raças de corte. O CMS para novilhos Holandeses foi 2 a 17% superior (média de 12%) do que o CMS de novilhos de raças de corte de pesos iniciais semelhantes (Garrett, 1971, Fox & Black, 1984, e Fox et al., 1992). O maior CMS de novilhos Holandeses pode ser devido à maior demanda energética para manutenção e crescimento – animais Holandeses parecem ter uma maior proporção de vísceras e trato digestivo, os quais aumentam sua taxa metabólica. O maior CMS de bovinos Holandeses também pode ser atribuído ao seu maior tamanho à maturidade (Hicks et al., 1990a).
Animais na fase de terminação apresentam menor capacidade de consumo de MS, em porcentagem do PV e em g/kg de PV0,75, que os animais durante a fase de recria. A composição corporal, especialmente a porcentagem de gordura corporal, parece afetar a ingestão de alimentos. À medida que o animal se aproxima da maturidade, mais gordura é depositada. Em geral, quanto mais gordo o animal, menor o consumo de alimentos, para qualquer tamanho corporal. Outra causa seria o efeito de retroalimentação negativa do tecido adiposo, através do hormônio leptina, no controle de consumo. No Brasil, pela preferência do mercado consumidor pela carne bovina mais magra, há a tradição de abater animais com menor grau de acabamento, fazendo com que este efeito depressor da gordura no consumo não seja tão pronunciado.
Particularmente em grandes confinamentos norte-americanos, algumas empresas já estão disponibilizando ferramentas eletrônicas que permitem monitorar a variabilidade diária no CMS. Este acompanhamento do consumo de alimentos pode ser utilizado na identificação de indivíduos ou grupos com problemas potenciais, a tempo de evitar perdas econômicas mais severas (Sowell et al., 1999). Além disso, nutricionistas e confinadores comumente atribuem distúrbios metabólicos, tais como a acidose sub clínica, ao comportamento alimentar errático dos bovinos. Esta percepção é baseada no fato de que a variabilidade no consumo de dietas com alta proporção de grãos comprometem a manutenção do pH ruminal em patamares suficientemente altos para uma adequada função ruminal (pH ruminal >5,6 a 5,8) (McAllister et al., 2002). Se piquetes ou currais com baixo CMS puderem ser detectados precocemente, no início do período de confinamento, medidas corretivas apropriadas podem ser adotadas. Entre estas medidas poderíamos citar: alteração do número de animais por piquete, alteração na proporção de forragem ou inclusão de forragem com diferente tamanho de partícula, adição de uma forragem de maior aceitabilidade ou com menos poeira na dieta, adição de aditivos como tilosina, ionóforos e tamponantes, alteração na forma de tratamento do grão, aumento do tamanho de cocho por animal, reparação de cochos ou bebedouros danificados, remoção de lama dos currais, manejo de enfermidades sub clínicas ou, finalmente, o descarte e venda de animais com desempenho já comprometidos. Tais medidas podem incrementar o consumo ou reduzir as perdas econômicas associadas com baixos CMS e ganhos diários (Hicks et al., 1990a, e Hicks et al., 1990b).
Hicks et al. (1990a) afirmaram que já que o CMS é a base pata cálculo ou predição das exigências nutricionais, das taxas de ganho e dos lucros, este consumo deve ser calculado com a maior acurácia possível. Do ponto de vista do confinador, consumo alimentar representa 70 a 80% do investimento e uma estimativa confiável deste investimento é crítica para o sucesso desta operação (DeHaan et al., 1995).
Muitos autores afirmavam no passado que o maior consumo de alimentos verificado pelas raças européias frente ao Nelore era o reflexo da intensa seleção para ganho de peso nas raças européias, já que incrementos de ganho de peso implicavam em maior consumo de alimentos. É importante acompanhar qual será o efeito da intensa seleção para ganho de peso que está ocorrendo na raça Nelore no consumo e na eficiência alimentar. Será que com a maior ênfase na seleção para ganho de peso na raça Nelore, haverá efeitos ou repercussões no consumo e na eficiência alimentar desta raça?
Referências bibliográficas
ALMEIDA, R. & D.P.D. LANNA. 2003. Influence of breed on performance and dry matter intake by feedlot bull calves in Brazil. J. Anim. Sci. 81, Suppl. 1 (aceito para publicação).
BEAVER, E.E., J.E. WILLAMS, S.J. MILLER, D.L. HANCOCK, S.M. HANNAH & D.L. O´CONNOR. 1989. Influence of breed and diet on growth, nutrient digestibility, body composition and plasma hormones of Brangus and Angus steers. J. Anim. Sci. 67:2415-2425.
BOYLES, S.L. & J.G. RILEY. 1991. Feedlot performance of Brahman x Angus versus Angus steers during cold weather. J. Anim. Sci. 69:2677-2684.
DeHAAN, K.A., M.T. VAN KOEVERING & M.L. GIBSON. 1995. The effect of age, background, and gender on feed intake by feedlot cattle. Symposium: Intake by feedlot cattle. Oklahoma Agric. Exp. Sta. Res. Rep. P-942:9-22.
FERRELL, C.L. & T.G. JENKINS. 1995. Body composition effects on maintenance, feed intake and efficiency. Symposium: Intake by feedlot cattle. Oklahoma Agric. Exp. Sta. Res. Rep. P-942:23-30.
FOX, D.G. & J.R. BLACK. 1984. A system for predicting body composition and performance of growing cattle. J. Anim. Sci. 58:725.
FOX, D.G., C.J. SNIFFEN, J.D. O´CONNOR, J.B. RUSSELL & P.J. VAN SOEST. 1992. A net carbohydrate and protein system for evaluating cattle diets: III. Cattle requirements and diet adequacy. J. Anim. Sci. 70:3578-3596.
FRISCH, J.E. & J.E. VERCOE. 1977. Food intake, eating rate, weight gains, metabolic rate and efficiency of feed utilization in Bos taurus and Bos indicus crossbred cattle. Anim. Prod. 25:343-358.
GARRETT, W.N. 1971. Energetic efficiency in beef and dairy steers. J. Anim. Sci. 32:451.
HICKS, R.B., F.N. OWENS, D.R. GILL, J.W. OLTJEN & R.P. LAKE. 1990a. Daily dry matter intake by feedlot cattle: Influence of breed and gender. J. Anim. Sci. 68:245-253.
HICKS, R.B., F.N. OWENS, D.R. GILL, J.W. OLTJEN & R.P. LAKE. 1990b. Dry matter intake by feedlot beef steers: Influence of initial weight, time on feed and season of year received in yard. J. Anim. Sci. 68:254-265.
HUFFMAN, R.D., S.E. WILLIAMS, D.D. HARGROVE, D.D. JOHNSON & T.T. MARSHALL. 1990. Effects of percentage Brahman and Angus breeding, age-season of feeding and slaughter end point on feedlot performance and carcass characteristics. J. Anim. Sci. 68:2243-2252.
KAPPEL, L.C., F.G. HEMBRY, P.E. HUMES, P.E. SCHILLING & R.H. KLETT. 1972. Climatic, breed and ration effects on feedlot performance and carcass characteristics of steers. J. Anim. Sci. 35: 591-597.
KARUE, C.N., J.L. EVANS & A.D. TILLMAN. 1972. Metabolism of nitrogen in Boran and in Hereford-Boran crossbred steers. J. Anim. Sci. 35:1025-1030.
KREHBIEL, C.R., K.K. KREIKEMEIER & C.L. FERREL. 2000. Influence of Bos indicus crossbreeding and cattle age on apparent utilization of a high-grain diet. J. Anim. Sci. 78:1641-1647.
LEDGER, H.P., A. ROGERSON & G.H. FREEMAN. 1970. Further studies on the voluntary food intake of Bos indicus and Bos taurus and crossbred cattle. Anim. Prod. 12:425-431.
McALLISTER, T.A., K.S. SCHWARTZKOPFGENSWEIN, K.E. BEAUCHEMIN, D.J. GIBB, M.N. STREETER, D.D. HICKMAN & D.H. CREWS. 2002. Metabolic consequences of feeding behavior and intake in feedlot cattle. J. Anim. Sci. 80, Suppl. 1: 112-113.
NATIONAL RESEARCH COUNCIL. 1996. Nutrient Requirements of Beef Cattle, Seventh Ed. Washington, D.C.: National Academy Press.
SOWELL, B.F., M.E. BRANINE, J.G.P. BOWMAN, M.E. HUBBERT, H.E. SHERWOOD & W. QUIMBY. 1999. Feeding and watering behavior of healthy and morbid steers in a commercial feedlot. J. Anim. Sci. 77:1105-1112.
VAN SOEST, P.J. 1994. Nutritional Ecology of the Ruminant. Second Edition. Cornell University Press.
___________________________
1Rodrigo de Almeida é Médico Veterinário, Professor UFPR e PUCPR, Doutorando ESALQ/USP
2Dante Pazzanese Lanna é Engenheiro Agrônomo, Professor ESALQ/USP, Ph.D.
